FMS
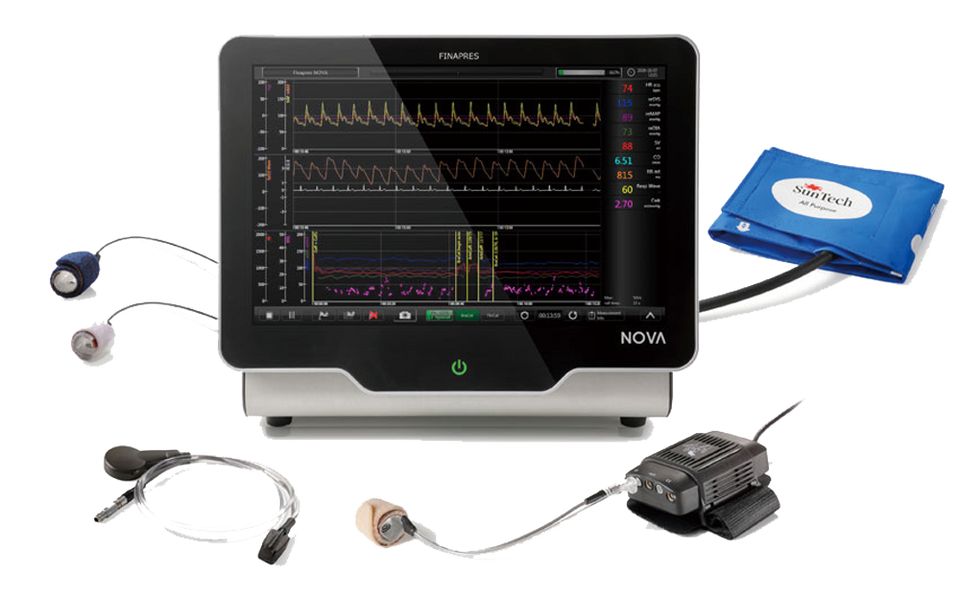
Finapres Medical Systems社 Finapres® NOVAは、革新的な血行動態モニタリングシステムです。
指用カフを使用するだけで、正確な非侵襲的連続血圧モニタリングが可能です。
このたび、ゼロシーセブン株式会社では FMS社 ホームページ を一新いたしました。
スマートフォンやタブレットでの表示にも対応しましたので、デバイスを問わずにいつでも閲覧いただけます。
つきましては、FMS社ホームページのアドレスを「お気に入り」や「ブックマーク」に登録されているお客様には、大変お手数をおかけいたしますが、設定の変更をお願いいたします。
■FMS社 ホームページURL
(旧):http://finapres.jp → (新):https://www.0c7.co.jp/products/FMS/
連続血圧・血行動態測定装置:Finapres® NOVA
Finapres® NOVAは、指先に専用のカフを装着するだけで、一拍毎の血圧の連続測定が行えるモニター一体型の装置です。
上腕カフを用いた血圧測定機能を備えており、上腕で計測した収縮期及び拡張期血圧値を指先計測値に反映させるキャリブレーション機能を有しています。
詳しく→